ホタル百科事典/ホタルの生態2-6.
![]()
ホタル幼虫の食べ物であるカワニナ(Semisulcospira libertina libertina)は、貝類の中では、腹足類(Gastropoda)に属します。カワニナ属は、日本には約40種が分布しています。それぞれの種の分布域は狭く、これらの内の14種が環境省のレッドデータブックに掲載されていますが、カワニナ属を分類すると大きく2つに分けることが出来ます。1つは、琵琶湖水系のみに生息する固有種群と、もう1つは全国に分布する広域生息種です。琵琶湖には、固有種も含めて以下の16種が生息しています。広域生息種においては、カワニナ類とその他のカワニナ属に分類できます。その他のカワニナ属とは、チリメンカワニナ、ヒタチチリメンカワニナ、クロダカワニナ等であり、これらとは違うものをカワニナ類と呼んでいますが、その種類は学者によってまちまちで3種〜6種と言われています。カワニナ属は、生息環境によって形態的特徴が異なっており、外見から種類を特定することが難しく、染色体数の違いまで調べなければ分からないこともあるようです。日本に生息するカワニナについては、2000年4月12日に環境省より陸・淡水産貝類のレッドリストが公表されています。
日本産カワニナ類
アマミカワニナ Stenomelania costellaris
ムチカワニナ Stenomelania cuenulatus
ヨシカワニナ Stenomelania juncea
スグカワニナ Stenomelania uniformis
ナカセコカワニナ Biwamelania nakasekoae
オガサワラカワニナ Stenomelania boninensis
イボアヤカワニナ Tarebia granifera
ホソマキカワニナ Biwamelania arenicola
クロカワニナ Biwamelania fuscata
モリカワニナ Biwamelania morii
イボカワニナ Biwamelania multigranosa
カゴメカワニナ Biwamelania reticulat
シライシカワニナ Biwamelania shiraishiensis
タケシマカワニナ Biwamelania takeshimaensis
クロダカワニナ Semisulcospira kurodai
フトマキカワニナ Biwamelania dialata
ナンゴウカワニナ Biwamelania fluvialis
オオウラカワニナ Biwamelania ourense
タテジワカワニナ Biwamelania rugosa
チリメンカワニナ Semisulcospira reiniana
ヒタチチリメンカワニナ Semisulcospira libertina hidachiensis
ヤマトカワニナ Semisulcospira (Biwamelania) niponica
ハベカワニナ Semisulcospira (Biwamelania) habei Davis
タテヒダカワニナ Semisulcospira (Biwamelania) decipiens
タケノコカワニナ Stenomelania costellris loebeckii
トウガタカワニナ Thiara (Pseudoplotia) scabra
ヌノメカワニナ Semisulcospira tuberculatus
オオカワニナ Semisulcospira libertina gigas
ミスジカワニナ Semisulcospira libertina japonica
キタノカワニナ Semisulcospira libertina dolorosa
琵琶湖のカワニナ固有種群

・ヤマトカワニナ Biwamelania
niponica
・ホソマキカワニナ Biwamelania arenicola
・タテヒダカワニナ Biwamelania
decipiens
・フトマキカワニナ Biwamelania
dialata
・ナンゴウカワニナ Biwamelania fluvialis
・クロカワニナ Biwamelania
fuscata
・ハベカワニナ Biwamelania habei
・モリカワニナ Biwamelania
morii
・イボカワニナ Biwamelania multigranosa
・ナカセコカワニナ Biwamelania
nakasekoae
・オオウラカワニナ Biwamelania ourense
・カゴメカワニナ Biwamelania
reticulata
・タテジワカワニナ Biwamelania
rugosa
・シライシカワニナ Biwamelania
shiraishiensis
・タケシマカワニナ Biwamelania
takeshimaensis
・チリメンカワニナ Semisulcospira reiniana
カワニナは、螺旋状に巻いた貝殻と蓋(operculum)を持っています。蓋は卵形で小旋型をしています。蓋は腹足類のみに形成され、巻貝の数少ない共通形質の1つです。体は頭足塊と内臓塊に分かれ、頭部には1対の頭部触角(cephalic tentacle)と眼が発達しています。頭部触角も腹足類のみに見られる構造です。そして触角の基部に眼を持っています。足は扁平で匍匐に適した形をしており、足の筋肉を波打たせながら移動します。この移動(這う)は、動物の中で一番消費エネルギーの多い方法であると言われています。また、貝殻を生成するにも多くのエネルギーが必要であると言われています。
カワニナは体の外側を覆う外骨格(exoskeleton)として貝殻を右巻(dextral)に形成しています。貝殻の主な機能は体の防御です。硬い殻の中に閉じこもってしまえば、捕食や環境の変化から逃れることができます。もうひとつの機能は体を支えることです。
貝殻は外套膜(がいとうまく)から分泌されます。外套膜は内臓塊全体を覆っていますが、主に貝殻の形成に関与する部分は外套膜の縁辺部の外套膜縁(mantle
margin)と呼ばれるところです。殻の形成には、まず殻皮(periostracum)と呼ばれる有機質の膜が分泌され、その膜の上に炭酸カルシウムの結晶が成長します。殻皮は殻が成長するための基質として作用します。貝殻は外套膜によって直接形成されるのではありません。外套膜内部で結晶が生産させて運ばれるのではなく、外套膜と貝殻の間を満たす液体の化学反応を通じて形成されています。この液体は外套膜外液(extrapallial
fluid)といい、貝殻を形成するメカニズムをバイオミネラリゼーション、生成した無機-有機複合体はバイオミネラルと呼ばれています。通常カルシウムCaは、カルシウムイオンCa2+といったイオンの形で水中に存在しますが、貝が持っている炭酸脱水酵素の働きによって水分子が二酸化炭素と反応して炭酸水素イオンHCO3-を生成し、それがカルシウムイオンと結合して炭酸カルシウムCaCO3という結晶となり貝殻を形成していくと考えられています。カワニナについての研究はされていませんが、モノアラガイやその他の貝では研究され解明されています。貝殻は均質な非晶質の塊ではなく、無数の微小な結晶の集合体です。貝殻の炭酸カルシウム(calcium
carbonate)にはカルサイト(calcite)とアラゴナイト(aragonite)の2つの結晶型があり、貝殻は複数の殻層(shell
layer)が重なり合って形成されています。貝殻の成長は殻の縁辺部に結晶が付加されながら大きくなっていきます。このような成長様式の特徴として、成長線(growth
line)が形成されます。成長の不連続面が1つ1つの成長線として殻に記録されます。また貝殻は螺旋状に極めて規則的に成長します。カワニナは、殻表面に多くの細かい筋があり、また時に2〜3の紫褐色帯が、殻口外唇の内側に現れることもあります。しかしながら、同じ種類でもその生息環境によって色、形、大きさに相違があります。カワニナは生息環境に適応する柔軟性をもっており、「表現型可塑性」を持っていると言えます。例えば、流れの緩やかな場所に生息しているカワニナは細長く、速い場所に生息するカワニナは、大きく「ずんぐりむっくり」型という形態的特徴があります。同じ河川にすむ同じ種類のカワニナでも、数十メートル離れただけで同じ種類とは思えないほど色や形が違っている場合もあります。貝殻の色については、餌となる藻類の色に影響を受けていると考えられています。貝殻は、有機質の殻皮(periostracum)に覆われていますが、殻皮は一度剥がれると修復できないため、形成後に長時間が経過した殻頂部では殻皮が剥がれて、殻の一部が溶食されてるカワニナもいます。
山口県秋芳洞口で石灰が沈着して、殻頂が完全に残るのは、スジマキカワニナ(S.B.nassaefounis
KURODA et KANAMARU)という形態型です。年間を通じて15℃という一定の水温と多量の石灰によって、殻が丈夫で形がずんぐりとした特殊な形態となっています。

同じ種類でも、それそれ生息している環境、水質によって殻の形や色が違っています。
流水の純淡水に生息する巻き貝はカワニナ属だけであり、北海道南部、本州、四国、九州、沖縄の各地に分布が見られます。
関東では、ヒタチチリメンカワニナが比較的多く見られます。
![]()
カワニナは、雌雄異体の卵胎生で、3cmを越える大きな母貝の体内には、孵化した稚貝がたくさん入っています。5月上旬から10月上旬にかけての5ヶ月間は、毎日2〜15個ほどの稚貝を生みます。ただし、夏期は春と秋に比べて少なくなるようです。(年間産仔数800〜2000個体)種類によっては、1回に約30個の稚貝を約1週間連続して産み、その後しばらくの間、稚貝の産出を休み、また、産みはじめるものもいるようです。稚貝を生み始めるには、水温の上昇や直射日光が体に当たることによる体温上昇が刺激になるようです。また、河川のどの場所でも産むのではなく、川岸の水草が茂っているような場所や浅瀬が適しているようです。
5月始めに生まれた0.2mmから0.5mmのカワニナの稚貝は、7月には2mmに育ち、これがゲンジボタルの孵化したばかりの幼虫の絶好の食べ物になるのです。カワニナは種類によって稚貝を産む数が異なっており、例えばチリメンカワニナは、稚貝が大きく産出数も少ないです。(年間産仔数100〜350個体)こういった種類はゲンジボタル幼虫の食べ物としては適していません。稚貝は、殻高約2cmに成熟するまで6〜7ヶ月かかり、翌年から稚貝を産み始めます。成長速度は、1年目が最も早く、産仔が開始される2年目以降は遅くなっていくようです。そして2〜3年で殻高は30mm、殻径13mmほどになり成長は止まります。カワニナは、ゲンジボタル幼虫の捕食原因以外により殻の直径が1mm大きくなるごとに60%ずつ死んでいき、1年後からは死亡率10%になり、2年後には3%しか生き残れないという報告もあります。カワニナの寿命はおよそ6年で、9年〜12年ほど生きるものもいるようです。
カワニナの食性を生息地における摂食物から判断してみると、カワニナはどの場所においても(摂食物の存在量の相違はあるものの)、付着藻類、落ち葉、その他植物、死んだ魚などを食べています。これは解剖した結果でもそれらを確認することができます。
付着藻類と言ってもたいへん多くの種類があり、その分類の仕方も様々です。カワニナと同じ場所に生息する付着藻類は、その生態的・形態的特徴から、着生藻類(periphyton)、岩表生藻類(epilithic algae)と堆積物表生藻類(epipelic algae)に大別できます。着生藻は、岩、砂粒、堆積物、植物体の他、他の藻類の表面にまで付着しています。この中には基質にしっかりと付着せず被っているようなものや、しっかりと仮根などで活着しているものなどがあります。類岩表生藻類は、比較的水流の強い細かい堆積物が積もらないところに生息しています。この仲間は、よく発達した付着機構をもち、ある程度の水流にも流されない形態を持っています。堆積物表生藻類は、比較的水流の弱い砂上、砂内部などに生息しています。
これら藻類は、珪藻類と藍藻類、緑藻類で構成されます。
珪藻類
珪藻類の大きな特徴は、体の側に殻をもっていることです。この殻の主成分は珪酸で珪藻類はその名前からも推察されるように、珪素と関係があります。珪素という元素(Si)は、珪酸(nSiO2
・xH2O)の形態で種々の生物中に存在していますが、その生理的な役割はあまりわかっていません。これに対して珪藻類は、自分自身の細胞質を珪酸で包んで保護しているという点で、ユニークな生物(植物)であると言えます。
そもそも珪素(Si)は,岩石圏の主要成分として二酸化珪素(SiO2),珪酸塩の形で多量に存在しています。 珪藻類は珪酸で細胞を包んでいるために、大きくなる種類がいません。大きなものでも0.5mm以下で、0.02mmくらいです。これは、珪藻類の殻がガラス質なので、柔軟性がなく、割れやすいからであると言われています。珪藻はこの割れやすい殻を保護するために、光合成で得られた糖類でゲルを作り、殻をその中に埋め込んでいるのです。
この殻は、動物が食べると簡単に消化されてしまいます。珪藻類は、英語圏ではダイアトムdiatom(s)と呼ばれています。di-はmono-、di-、tri-、tetra−、penta-のdi-で数字の2を意味しています。atomは原子(つまり分割できないもの)という意味ですから、珪藻diatomは、
"分割できない2つの部分から成る" という意味を持っています。珪藻類は[ ]
という形の殻からできているので、分解するとカプセルの内側と外側にバラけてしまいます。日本語では成分に着目して「珪藻」、英語では形に着目してdiatomと呼ばれています。
珪藻は、緑色の葉緑素の他に赤茶色のカロチノイド色素、キサントフィル、フコキサンチン,ディアディノキサンチン,ディアトキサンチンなどの黄色をした補助色素も含まれているので,黄褐色系統の色をしています。
珪藻が光合成の結果として作り出す有機貯蔵物質は,緑色植物にみられるデンプンとは違い,クリソラミナリン(β−1,3グルカンを主体)とよばれる多糖類および油脂です。油の成分は、種によって,またその生育環境によって組成が異なりますが,代表的なものは不飽和脂肪酸のC14のミリスチン酸,C16のパルミチン酸やパルミトレイン酸,
C18のオレイン酸,C20のEPA(イコサペンタエン酸)であり、C20のEPAが豊富であることが特徴です。
カワニナと同じ環境に生息するものとしては、ハリケイソウ、
アクナンテイス、クチビルケイソウ、クサビケイソウなどがいます。
藍藻類
藍藻類は原核生物としての細胞構造持った唯一の藻類です。藍藻類は、世界中至る所に分布し、温度に関係なく湿っていれば海水でも淡水でも、また空気に露出した所でも生育しています。水中では、深い所より浅い場所が適しているようで、例えば水たまりや滝の岩石の表面、温泉の周囲などに多く見られます。藍藻類には空気中の窒素を固定する能力(NH4~+を吸収してアミノ酸を合成する)があるため、植物の根毛周囲にも多く見られます。また、藍藻類には、石灰を溶かす能力のあるものや逆に石灰を析出するものもあり、石灰土中にも多く繁茂しています。藍藻類の中で最大のグループは2つの亜目に分けられています。一つはネンジュモ亜目でヘテロシスト=異形細胞(他の細胞と違った形をしており、窒素固定能力を持つ)を持っており、もう一つはユレモ亜目でヘテロシストは持っていません。中にはアキネート(休眠細胞)を作り出して低温や乾燥に耐えることができものもあり、このため、過酷な環境下でも生息できます。このグループは糸状の形をしており、分裂によって増殖します。
カワニナと同じ環境に生息するものとしては、クロオコックス、ホメオスリックス、コンボウランソウ、ユレモなどがいます。
緑藻類
単細胞性のもの、群体を形成するもの、肉眼的な藻体を形成するものと体制はさまざまで、非常に変化に富んでいます。
細胞内に葉緑体をもち、光合成をしています。葉緑体は陸上植物と同じ光合成色素をもち、鮮やかな緑色です。
しばしば葉緑体内にピレノイドとよばれる球状の構造物があります。ボルボックス目以外のものは細胞に繊毛や鞭毛などの運動性の毛がないため活発に遊泳することはありません。大多数は全く動きません。細胞壁は、セルロース(cellulose)で出来ています。セルロース(cellulose)とは、分子式 (C6H10O5)n の炭水化物(多糖類)です。植物細胞の細胞壁および繊維の主成分で、天然の植物質の1/3を占め、地球上で最も多く存在する炭水化物となっています。セルロースはブドウ糖が鎖状につながったものですが、セルロースそのものを分解して栄養源としているものはシロアリや微生物であり、ほとんどの動物(牛などの反芻動物では胃のなかにいる微生物がセルロースを糖に分解している)では、食物繊維として分解されずに排出されます。
カワニナと同じ環境に生息するものとしては、キヌミドロ(スティゲオクロニウムStigeoclonium
spp.)がいます。
カワニナの成長や繁殖には、他の生物同様に三大栄養素(炭水化物、脂質、タンパク質)と無機質とビタミン類が必要ですが、以上の摂食物からは、炭水化物と脂質については十分に摂取できます。特に、大量のエネルギーを必要とするカワニナにとっては、珪藻類はもっとも重要な摂食物となっていると考えられます。それは、カワニナの貝殻の色からも推察できます。しかし、これではタンパク質はあまり摂取できません。魚の死骸等には、タンパク質が豊富ですが、頻繁に食べることができるほど供給はされません。また、落葉や腐りかけた葉や茎、その他植物自体にもタンパク質はそれ程多くは含まれていません。カワニナは、「低タンパク高カロリー」が食性の特徴なのでしょうか?
実は、一般的に淡水性の貝類は、微生物も栄養源として摂取していることが知られています。これらは、消化が速いため胃の中からは見つけることが出来ませんが、カワニナにおいても摂取している可能性は十分考えられます。この微生物は、水中の落葉や水中で腐りかけた茎葉、魚の死骸等、動植物の遺体等のような体組織由来の物質、排泄物といった生物由来の物体(デトリタス)を栄養源として繁殖します。
デトリタスは、特に流れの緩やかな部分に沈殿します。このデトリタスには多くの微生物が繁殖し、タンパク質や低分子の糖類、脂質などは速やかに分解吸収されます。難分解性の多糖類においても、繁殖した微生物はデトリタスから炭素を、環境水中から窒素やリンを吸収(窒素同化してアミノ酸を合成)し、自らの細胞構成物質を合成します。つまり、微生物が繁殖したデトリタスを摂食することによりタンパク質に富んだ微生物も一緒に吸収することになります。
カワニナの繁殖や稚貝の生育には、タンパク質が重要であるという報告があります。飼育しているカワニナにタンパク質を多く含む餌を与えると稚貝を沢山産み、また稚貝の生育も良いのです。自然界では、カワニナの繁殖域と稚貝の生育域は河川の浅瀬や岸辺の水草が茂っている部分ですが、そこにはデトリタスが堆積もしくは腐食植物が多く存在し、実際にそれらを食べています。つまりカワニナの親貝や稚貝は、落葉や腐りかけた草を食べることによって、そこに繁殖している微生物も摂食してタンパク質を得ていると考えられるのです。
カワニナは集団生活者であり、生息環境が整っていれば、特に西日本では群落をつくって生活しています。カワニナは、殻の成分が炭酸カルシウムですのでカルシウムイオンを多く含む水を好みます。石灰石が多くある川や上流に鍾乳洞がある河川などでは、たくさんのカワニナを見ることがしばしばあります。
カワニナの生息条件としては、カワニナは呼吸量が多いために、溶存酸素量は、飽和に近く、水深が15から30センチくらいの川が好適です。水温については、高水温には弱く、25℃を大幅に超えると死んでしまいます。また、年間を通じて低水温ですと、貝の成長が悪かったり、子産数が少ない或いは、子産期間が短いという影響があるようです。流速はそれほど関係ないようです。ほとんど流れのないところや人がやっと立っていられるほどの流れの速いところにも生息しています。ただし、流速1m/secになると流されてしまうようです。また、底質は礫から泥状までと多様です。また、冬期になると浅瀬に集まり川底の石の下や泥の中などに潜ってじっとしているものや水温が2℃でも活発に活動しているカワニナもいます。ただし、これらはそれぞれの環境に適応して生息しているのであり、ある生息地のカワニナを採取して別の環境に放流しても生息できない場合もあります。例えば、カワニナは遡上行動を行いますが、流れが穏やかで浅い場所に生息するカワニナを流速の早い場所に移動させると遡上はせずに、流れが穏やかで浅い場所まで下流へ下っていきます。
β−中腐水性水域 |
貧腐水性水域 |
|
|---|---|---|
溶存酸素 |
かなり多い |
多い |
BOD |
2.5〜5ppm |
2.5ppm以下 |
硫化水素の発生 |
なし |
なし |
水中の有機物 |
脂肪酸のアンモニア化合物が多い |
有機物は分解されてしまっている |
水中の細菌数 |
10万以下/1ml |
100以下/1ml |
原生動物 |
太陽虫や吸管虫類の汚濁に弱い種類,渦鞭毛虫類が出現 |
鞭毛虫類,繊毛虫類が少数出現 |
水生動物 |
淡水海綿,こけ虫類,ヒドラが出現,貝類,小型甲殻類,昆虫の幼虫,両生類,魚類の多くの種類が出現 |
昆虫の幼虫の種類が多い,他に各種の動物が出現 |
水生植物 |
珪藻,緑藻,接合藻の多くの種類が出現,ツヅミモ類はここが主要な分布域 |
水中の浮遊藻類は少ないが,着生藻類は多い |
カワニナの外敵としては、サワガニ、ハバヒロビル、クマネズミ等が挙げられます。
また、寄生虫は約40種類いると言われていますが、人に害はないようです。
現在、カワニナはホタルの餌としか見られていません。カワニナ属に関する研究事例も少なく、研究といってもその養殖方法の探索に終始しています。ホタルの保護活動が盛んになり、自然河川にホタルを増やすためだけにある種のカワニナをばらまくことや、人工河川に業者から購入したある種のカワニナを投入し続けるという行為が普通に行われています。カワニナ属全般の生態についての研究が進まなければ、こうした行為も止まることはないかも知れません。自然生態系の中では、カワニナはホタル幼虫に食べられるだけの存在ではないのです。
 カワニナの幼貝
カワニナの幼貝
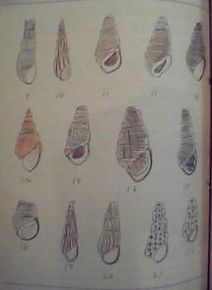 様々な種類のカワニナ
様々な種類のカワニナ
参考:カワニナの飼育と方法/カワニナの個体群動態とゲンジボタル生息との関係について